


Abstract
The present study defined age differences in several aspects of nicotine dependence using male mice of two age groups [postnatal day (PND) 28 and PND 70]. Adolescent and adult mice displayed differences in acute sensitivity to nicotine, rewarding and withdrawal effects, development of tolerance to nicotine, and nicotinic receptor function. In the condition place preference model, adolescent mice displayed a higher sensitivity to nicotine than adults. In addition, in spontaneous and mecamylamine-precipitated withdrawal models, adolescent mice displayed fewer withdrawal signs than adults. In response to acute nicotine, it was found that adolescent mice displayed greater nicotine-induced antinociception compared with adult counterparts in the tail-flick test. Furthermore, differences in tolerance to nicotine were also noted in that adolescents developed a significantly higher degree of tolerance to nicotine in the hot-plate test compared with adults. Finally, using rubidium efflux assays, it was found that adolescent nicotinic receptors in different brain areas displayed significantly increased functionality compared with adult receptors. These data indicate that the underlying receptor mechanisms of nicotine dependence differ for adults and adolescents, suggesting that the effectiveness of smoking cessation therapies will differ for various age groups.
More than 90% of adult smokers report their first use of tobacco before age 18 (Chassin et al., 1990). Moreover, the commencement of smoking at a young age is thought to increase addiction and to decrease the probability of successful cessation (Chassin et al., 1990; Chen and Millar, 1998; Colby et al., 2000; Kandel and Chen, 2000).
Nicotine is the main active ingredient in tobacco smoke that causes and maintains tobacco addiction. Abundant clinical and experimental data have revealed that nicotine produces tolerance and leads to psychological and physical dependence in adults. Moreover, nicotine causes molecular and cellular changes in the brains of adults. However, it is not known whether similar behavioral and neurobiological changes occur in adolescence. Because considerable neurodevelopment occurs during adolescence (Spear, 2000; Sowell et al., 2001; Giedd, 2004), nicotine may differentially affect the brains of adolescents compared with adults. Overall, studies conducted to date suggest that the rewarding and reinforcing effects of nicotine are enhanced in adolescent versus adult rodents. It has been shown that female adolescent rats that initiate nicotine self-administration during adolescence exhibit higher nicotine intake as adults relative to rats that initiate self-administration as adults (Levin et al., 2003). Adriani et al. (2002) showed that early adolescent mice displayed a marked preference for nicotine in drinking water, whereas middle and late adolescents showed no preference or avoidance, respectively. They also demonstrated that nicotine self-administration is increased in rats that received nicotine injections during adolescence (Adriani et al., 2003). In addition, Klein et al. (2004) showed that adolescent mice of both sexes would voluntarily consume nicotine orally in a dose-dependent manner. Prior studies (Risinger and Oakes, 1995; Martin and Itzhak, 2000) have found that nicotine produces a conditioned place preference in the adult mouse, but studies examining differences in age groups have yet to be conducted. In a rat model, Vastola et al. (2002) reported that in the conditioned place preference paradigm only animals conditioned during adolescence showed preference to nicotine. Likewise, Belluzzi et al. (2004) reported that animals exposed to nicotine during early adolescence (PND 28) showed preference, whereas older adolescents and adults failed to do so. The present investigation determined whether such age differences are also present in the mouse. In addition to enhanced rewarding effects in adolescent rodents, recent studies demonstrate that this age group may be less susceptible to withdrawal symptoms than adults. O'Dell et al. (2006) found that adolescents display fewer somatic signs than adults in several different dosing protocols. Furthermore, in a conditioned taste avoidance model, Shram et al. (2006) showed that adult rats display a dose-dependent avoidance to saccharin solutions paired with nicotine, whereas adolescents do not develop this avoidance. Behavioral differences in rewarding properties and withdrawal from nicotine may cause an increased vulnerability to nicotine addiction in adolescent animals. Although the mechanisms underlying these age differences in nicotine reward and withdrawal are yet to be elucidated, they can be attributed to various factors, such as differences in pharmacokinetics and pharmacological mechanisms, neurobiochemical changes, hormonal patterns, genetics, and environmental influences.
Because behavioral effects of nicotine may vary with the paradigm used, it is important to conduct studies where various behavioral responses are evaluated in the same setting and in parallel. Herein, we report on an exhaustive study investigating age differences in male mice. Specifically, the following questions were addressed to examine possible underlying differences in nicotine reward and withdrawal: 1) Do differential acute in vivo and in vitro sensitivities to nicotine between adolescent and adult animals exist? 2) If so, can these age-related differences in acute sensitivity explain differences in the development of tolerance to nicotine following chronic exposure? and 3) If so, can these age-related differences in acute sensitivity explain differences in nicotine dependence behaviors such as reward and withdrawal?
For that, nicotine dose-response curves in the conditioned place preference test in early adolescent (PND 28) and adult (PND 70+) mice were determined. Furthermore, differences in somatic and affective withdrawal signs were investigated in these two age groups. We then compared the potency of the effect of nicotine after acute administration through a series of pharmacological responses such as antinociception, hypothermia, and locomotor activity. Next, differences in the degree of tolerance to nicotine after chronic exposure were examined between adult and adolescent mice using the same pharmacological responses mentioned above. Finally, a biochemical assay that measures nicotine-stimulated 86Rb+ efflux in synaptosomes isolated from mouse brain regions was used to compare nAChR functionality between adolescents and adults.
Materials and Methods
Animals
Experimentally naive male ICR mice were obtained from Harlan (Indianapolis, IN). They were housed either five or six per cage depending on the experiment, and they were given free access to food and water. For all studies, early adolescent mice arrived on PND 21, and adult mice arrived on PND 70. Animals were allowed to acclimate, and they were handled for 5 to 7 days before the start of all experiments, since it has been shown that this technique is important in reducing levels of stress that might affect experimental results (Grabus et al., 2006). Adolescent and adult mice were therefore between the age range of PND 28 to 36 and PND 75 to 84, respectively, while experiments were conducted. Animals were maintained in an American Association for Accreditation of Laboratory Animal Care approved facility and the study was approved by the Institutional Animal Care and Use Committee of Virginia Commonwealth University.
Drugs
(-)-Nicotine bitartrate was purchased from Sigma-Aldrich (St. Louis, MO). Mecamylamine hydrochloride was a gift from Merck Research Labs (West Point, PA). All doses are expressed as free base.
Acute Studies
Mice were given s.c. injections of nicotine, and they were tested in four pharmacological tests: analgesia (two assays, tail-flick and hot-plate), locomotor activity, and hypothermia.
Tail-Flick Test. Spinal antinociception was assessed by the tail-flick method of D'Amour and Smith (1941). In brief, mice were lightly restrained while a radiant heat source was directed onto the upper portion of the tail. A control response (2-4 s) was determined for each mouse before treatment, and test latency was determined after drug administration. To minimize tissue damage, a maximum latency of 10 s was imposed. Antinociceptive response was calculated as percentage of maximum possible effect (%MPE), where %MPE = [(test - control)/(10 - control)] × 100. The mice were tested 5 min after injection of nicotine.
Hot-Plate Test. Supraspinal antinociception was assessed by the hot-plate method. Briefly, each mouse was injected s.c. with nicotine, and it was tested 5 min after injection. Mice were placed on a hot-plate (Thermostat Apparatus, Columbus, OH) maintained at 55°C. Latency to reaction time (jumping or paw licking) was recorded. A control response (8-12 s) was determined for each mouse before treatment, and test latency was determined after drug administration. A maximum latency of 40 s was imposed. Antinociceptive response was calculated as %MPE, where %MPE = [(test - control)/(40 - control) × 100].
Locomotor Activity. Mice were placed into individual photocell activity cages (28 × 16.5 cm; Omnitech, Columbus, OH) 5 min after s.c. administration of nicotine. Interruptions of the photocell beams (two banks of eight cells each) were then recorded for the next 10 min. Data are expressed as number of photocell interruptions.
Body Temperature. Rectal temperature was measured by a thermistor probe (inserted 24 mm) and digital thermometer (YSI Inc., Yellow Springs, OH). Readings were taken just before and at 30 min after the s.c. injection of nicotine. The difference in rectal temperature before and after treatment was calculated for each mouse. The ambient temperature of the laboratory varied from 21-24°C from day to day.
Tolerance Studies
Mice were implanted with Alzet osmotic minipumps (model 2002; Alza, Palo Alto, CA) filled with either (-)-nicotine (48 mg/kg/day) or sterile physiological saline solutions. The minipumps were surgically implanted s.c. under sterile conditions with pentobarbital anesthesia (50 mg/kg i.p.). An incision was made in the back of the animals, and a pump was inserted. Animals were sutured and allowed to recover before being returned to their home cages. Mice were infused with nicotine and saline for 10 days, and on day 11, they were challenged with different nicotine doses and tested for antinociception (tail-flick and hot-plate tests) and hypothermia.
Nicotine-Induced Conditioned Place Preference
Place conditioning boxes consisted of two distinct sides (20 × 20 × 20 cm). A partition separated the two sides with an opening that allowed access to either side of the chamber, and this partition could be closed off for pairing days.
Handling Habituation. On Wednesday to Friday of the week before the start of the place conditioning procedure, mice in the conditioned place preference studies were handled once per day for approximately 2 min each. Handling experience plays an important role in the ability of nicotine to produce a conditioned place preference (Grabus et al., 2006).
Preconditioning Phase. On day 1, animals were placed in the boxes, and they were allowed to roam freely from side to side for 15 min, and time spent in each side was recorded. These data were used to separate the animals into groups of approximately equal bias.
Conditioning Phase. Animals were paired for 20 min with the saline group receiving saline in both sides of the boxes and with the drug groups receiving nicotine (0.05, 0.1, 0.5, 0.7, or 1 mg/kg) on one of the sides and saline on the opposite side. Drug-paired sides were randomized among all groups. Conditioning lasted for 3 days, with animals in the drug group receiving drug each day.
Test Phase. On the test day, no injections were given. Time spent on each side was recorded, and data were expressed as time spent on drug-paired side minus time spent on saline-paired side. A positive number indicated a preference for the drug-paired side, whereas a negative number indicated an aversion to the drug-paired side. A number at or near zero indicated no preference for either side.
Mecamylamine-Precipitated Withdrawal Studies
Naive mice were implanted with osmotic minipumps filled with either saline or nicotine (48 mg/kg/day) on day 1 (as described in “Tolerance Studies”). On the morning of day 8, mice were injected s.c. with 2.0 mg/kg mecamylamine, a nicotinic antagonist. Ten minutes following injection with the antagonist, mice were tested for withdrawal signs in the following manner: 5 min for anxiety-like behavior (on the elevated plus maze described below), 20-min observation of somatic signs (paw tremors, head shakes, backing, body tremors, and ptosis), hyperalgesia (via hot-plate test described above), and 30 min in locomotor activity chambers (described above).
The elevated plus maze is an apparatus consisting of two closed arms and two open arms. The mouse is placed in the center of the maze, and it is allowed to roam freely between the open and the closed arms for 5 min. The number of seconds the mouse spends in the open arms is counted by a counting device attached to beams located on both arms and in the middle of the plus maze. Scores are based on time spent in the open arms as an indication of anxiety-like behavior.
Spontaneous Withdrawal Studies
Naive mice were implanted with osmotic minipumps filled with either saline or nicotine (48 mg/kg/day) for 7 days (as described in “Tolerance Studies”). On day 8, mice were lightly anesthetized using ether, and minipumps were removed. A small incision was made on the back of the neck to remove the minipump, and the wound was closed with a suture. Twelve hours following removal of the minipump, mice were evaluated with the same withdrawal measures described above.
Repeated Injection Withdrawal Model
Naive mice were injected s.c. with 2.0 mg/kg nicotine three times a day for 4 days (injections were given at 8:00 AM, 12:00 PM, and 4:00 PM). On the morning of day 5, mice received 2.0 mg/kg mecamylamine s.c., and they were evaluated 10 min later in the same withdrawal measures described previously.
Rubidium Efflux Studies
Unless otherwise noted, all reagents were purchased from Sigma-Aldrich). Mice were rapidly decapitated, and four brain regions were dissected for use in the assay (striatum, frontal cortex, hippocampus, and thalamus). Synaptosomes were prepared according to Marks et al. (1993a,b). In brief, synaptosomes were prepared by hand homogenizing tissue in radioactive 0.32 M sucrose (1 ml/g tissue). After centrifugation, pellets were resuspended in radioactive load buffer (140 mM NaCl, 1.5 mM KCl, 2 mM CaCl2, 1 mM MgSO4,25mM HEPES hemisodium salt, and 20 mM glucose, pH 7.4). A 25-μl aliquot of the synaptosome suspension was incubated for 40 min with 10 μl of load buffer containing approximately 4 μCi of 86RbCl (PerkinElmer Life and Analytical Sciences, Boston, MA). After the synaptosomes were filtered onto glass fiber filters under gentle vacuum, the filters were rinsed with 0.5 ml of load buffer, and they were placed on the perfusion apparatus for washing with perfusion buffer (135 mM NaCl, 5 mM CsCl, 1 mM MgSO4, 2 mM CaCl2, 1.5 mM KCl, 1 g/l bovine serum albumin, 50 nM tetrodotoxin, and 25 mM HEPES hemisodium salt, pH 7.4) for 6 min. The filter containing synaptosomes was subsequently perfused continuously. Filters were stimulated for 1 min with various concentrations of nicotine (1, 5, 10, 20, 30, 50, 100, 200, or 300 μM) prepared in perfusion buffer followed by a 3-min wash with perfusion buffer alone. Twelve-second fractions were collected in 12- × 75-mm test tubes beginning 6 min into the perfusion. Samples were counted for 1 min each in a Wallac Wizard 3′ 1480 automatic gamma counter; (PerkinElmer Life and Analytical Sciences). The magnitude of 86Rb+ efflux response was calculated based on the increase in counts above baseline after stimulation of the tissue with nicotine. Data were calculated as fractional release (cpm/total cpm loaded on filter) for each fraction collected. The baseline was calculated for each mouse by fitting to an exponential equation the fractional release in fractions immediately preceding and following the peak. The area under the curve was calculated for each mouse using this mathematically derived baseline and the fractional release values in the peak.
Statistical Analysis
Statistical analysis of all behavioral studies was performed with mixed factor ANOVA with post hoc Tukey's test when appropriate. p values <0.05 were considered to be statistically significant. For chronic tolerance studies, effective dose 50% (ED50) values were calculated with 95% confidence intervals by unweighted least-squares linear regression as described by Tallarida and Murray (1987). Tests for parallelism were calculated according to the method of Tallarida and Murray (1987). If confidence limit values did not overlap, then the shift in the dose-response curve was considered significant. Potency ratios were also calculated to determine whether tolerance differences were significant between age groups. Nicotine stimulated 86Rb+ efflux was analyzed with a two-way ANOVA and a one-way ANOVA as a function of age. These were followed by Tukey post hoc tests. Effective concentration 50% (EC50) values were calculated by unweighted least-squares linear regression as described by Tallarida and Murray (1987). A p value <0.05 was considered statistically significant.

Dose-response relationship for nicotine-induced condition place preference in mice. Male mice of both adult (PND 75) and adolescent (PND 28) ages were injected s.c. with various doses of either saline or nicotine. Each point represents the mean ± S.E.M. of eight to nine mice. *, p < 0.05 from saline group.
Results
Nicotine-Induced Conditioned Place Preference. As expected, adult and early adolescent animals injected with saline in both chambers, showed no preference for either chamber. Compared with saline controls, male adult mice conditioned with the intermediate dose of 0.5 mg/kg nicotine showed a significant place preference (F5,51 = 2.603; p < 0.02) (Fig. 1). There were no significant preferences seen with lower doses of 0.05 or 0.1 mg/kg, and the effect disappeared at the higher doses of 0.7 and 1.0 mg/kg nicotine. In contrast, nicotine showed a significant place preference in adolescent mice at low doses of nicotine (0.05 and 0.1 mg/kg) as well as at the 0.5-mg/kg dose (F5,50 = 3.303; p < 0.01). Similar to the adult mice, the effect disappeared at the higher doses of 0.7 and 1.0 mg/kg nicotine.

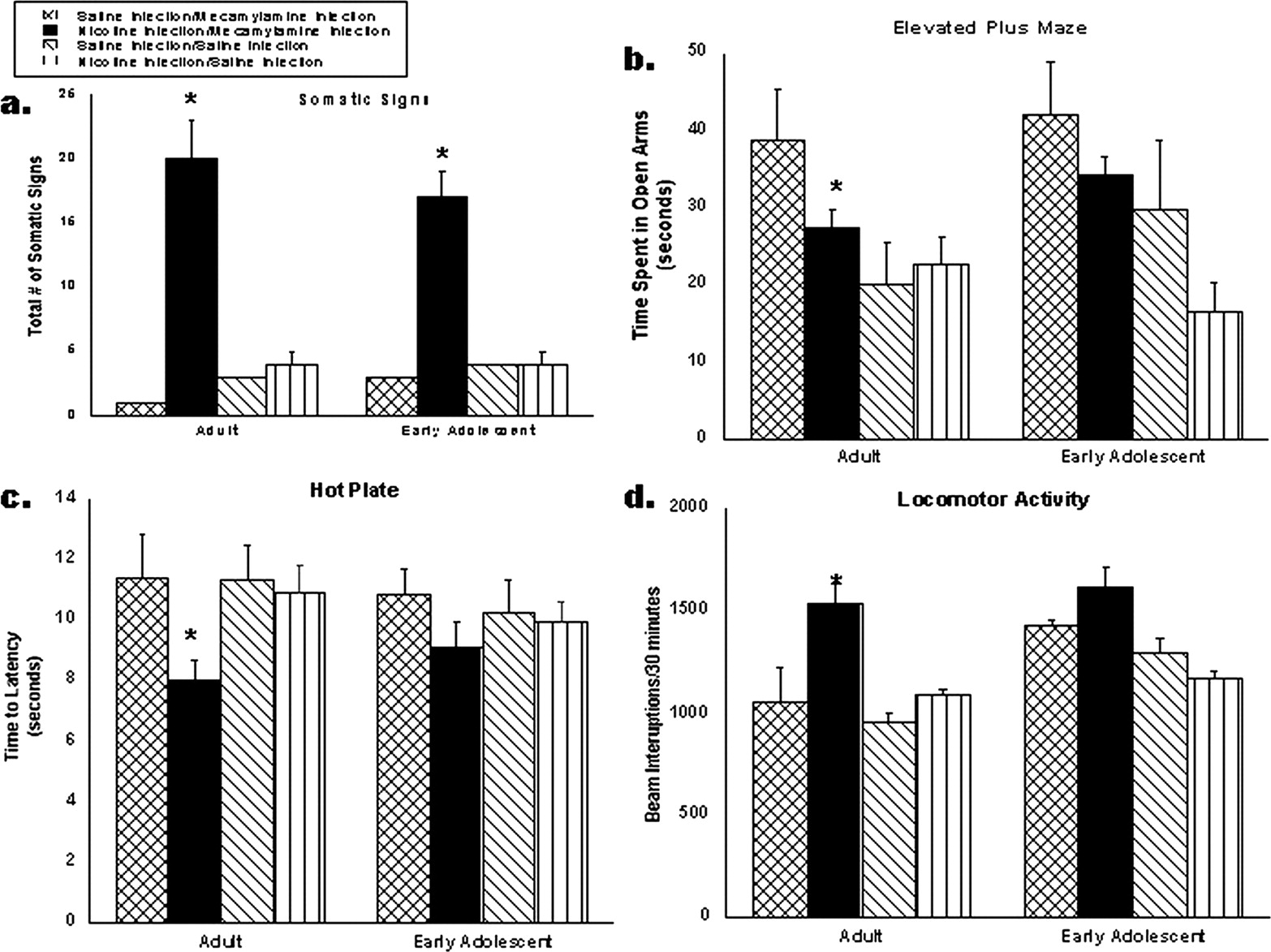
Mecamylamine-precipitated withdrawal in adult and adolescent mice. Adolescent (PND 28) and adult (PND 75) mice were chronically infused with nicotine at 48 mg/kg/day or saline for 7 days. On day 8, mice were injected with 2.0 mg/kg mecamylamine or saline s.c. to precipitate withdrawal, and they were evaluated in four tests: somatic signs (a), elevated plus maze (b), hot-plate analgesic test (c), and locomotor activity (d) (n = 10/group). *, p < 0.05 from saline group and #, p < 0.05 from adult nicotine treatment. Each point represents the mean ± S.E. of 12 mice. MP, minipump.

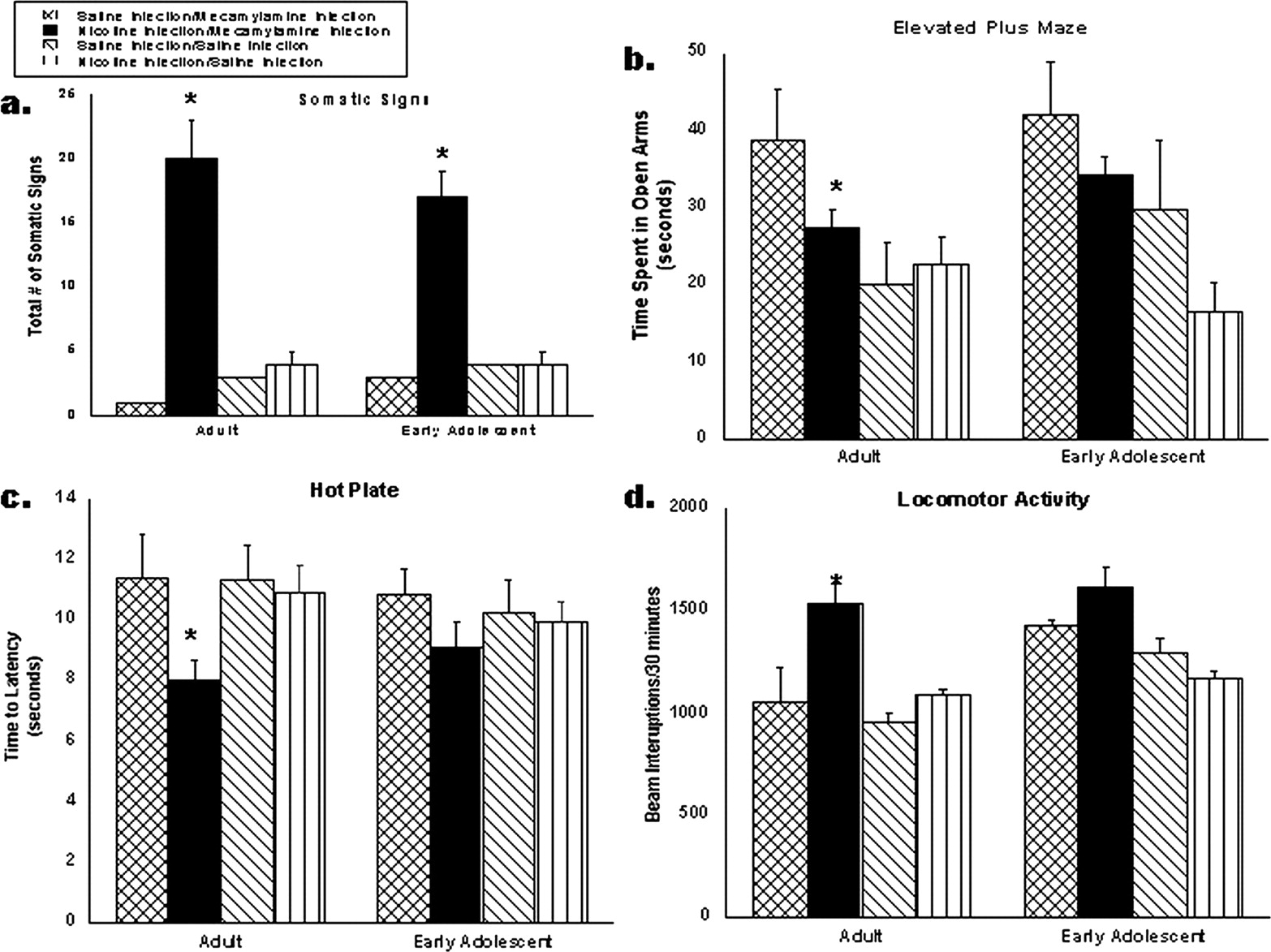
Spontaneous withdrawal in adult (PND 75) and adolescent (PND 28) mice. Male mice were chronically infused with nicotine at 48 mg/kg/day (dark bars) or saline (cross-hatched bars) for 7 days. Twelve hours after minipumps were removed, mice were tested for somatic signs (a), elevated plus maze (b), hot-plate (c), and locomotor activity (d). Each point represents the mean ± S.E. of 12 mice. *, p < 0.05 from saline group and #, p < 0.05 from adult nicotine treatment.
Precipitated Withdrawal Model. Measures of precipitated withdrawal in adult and adolescent mice are shown in Fig. 2. Two types of withdrawal signs were evaluated: physical (somatic signs, hyperactivity, and hyperalgesia) and affective (anxiety-like behavior in the plus maze). Adolescent mice displayed fewer withdrawal signs in all four measures compared with adults. Adolescents displayed lower number of somatic signs (Fig. 2a) and less hyperactivity in the locomotor chambers (Fig. 2d) compared with adults. In the elevated plus maze (Fig. 2b), only adults displayed withdrawal as indicated by a significant decrease in time spent in the open arms of the maze, an indication of anxiety-like behavior. Adolescents did not show withdrawal in this test. Likewise, in the hot-plate test (Fig. 2c), only adult mice displayed hyperalgesia.
Spontaneous Withdrawal Model. In the spontaneous withdrawal model, minipumps were removed on day 8, and no drugs were used to elicit withdrawal behaviors. As shown in Fig. 3, both adult and adolescent mice displayed a significant increase in the number of somatic signs after chronic treatment with nicotine; however, adolescent mice had significantly fewer of these signs compared with adults (Fig. 3a). Only adult mice displayed affective withdrawal signs in the spontaneous model as shown by the decrease in time spent in the open arms of the elevated plus maze (Fig. 3b). Results for the hot-plate (Fig. 3c) and locomotor activity tests (Fig. 3d) were similar to those of somatic signs in which both age groups displayed withdrawal symptoms, but adolescent withdrawal was significantly less than that of the adult mice. This confirmed the data from the precipitated withdrawal model.
Repeated Injection Withdrawal Model. The repeated injection model confirmed our previous withdrawal results with minipumps as shown in Fig. 4. Hyperactivity is only demonstrated by adult mice in the locomotor activity test (Fig. 4a). Furthermore, in the hot-plate test (Fig. 4b), adult mice displayed hyperalgesia, whereas adolescent mice failed to exhibit this effect. Only adolescent mice show anxiety-like behavior in the elevated plus maze (Fig. 4b). In somatic signs (Fig. 4a), both ages display withdrawal, but there is a trend for a decreased number of somatic signs in adolescents.
Pharmacological Effects of Nicotine after Acute Injection. To identify possible underlying differences observed in adolescent and adult mice in nicotine reward and withdrawal, we first explored the possibility of age differences in the sensitivity to acute nicotine injections. Dose-response relationships were established for nicotine in male mice of both ages by measuring antinociception (two tests), hypothermia, and hypomotility at the time of maximal effect (Fig. 5) and ED50 values (confidence limits; CL) values were then determined for each age in different tests (Table 1). Baseline levels in the tail-flick and hot-plate tests were not significantly different between the two ages, respectively (PND 28: 2.9 ± 0.2 and 12.5 ± 1.3 s; PND 75: 2.7 ± 0.3 and 13.0 ± 1.8 s).
Summary of nicotine potency in young and adult mice after acute injections Mice from two age groups (PND 28 and PND 75) were injected s.c. with various doses of nicotine and tested in the following responses: tail-flick test, hot-plate test, hypothermia, and locomotor activity. ED50 values (± CL) were calculated from the dose-response curve of the respective treatment, and they are expressed as milligrams per kilogram. Each dose group included 8 to 10 animals.
In the tail-flick test, early adolescent male mice displayed decreased sensitivity compared with male adult mice (Fig. 5a). ED50 values with confidence limits were 1.7 (1.3-3.6) and 1.0 (0.6-1.2) mg/kg for adolescent and adult age groups, respectively (Table 1). However, no significant differences between the two age groups were observed in the hot-plate, hypothermia, and hypomotility tests (Fig. 5, b-d). In addition, all acute responses to nicotine in adult and adolescent mice were blocked by mecamylamine at 1.0 mg/kg (data not shown).
Development of Tolerance to Nicotine after Chronic Exposure. As shown in Fig. 6, tolerance developed to the antinociceptive and hypothermic effects of nicotine in both adults and adolescents as reflected by the rightward shift in the dose-response curves. Furthermore, these shifts were significant as demonstrated by the significant increase (with nonoverlapping confidence limits) in ED50 values after chronic nicotine (Table 2). To determine whether the degree of tolerance was significantly different between the age groups, we calculated potency ratios for each group (Table 3). Adolescent male mice showed a higher degree of tolerance in the hot-plate test [potency ratios with confidence limits for adolescent and adults are 2.31 (2.03-2.62) and 1.75 (1.47-1.98), respectively]; however, tolerance to the tail-flick and body temperature developed at the same degree.
ED50 values of tolerance studies after chronic administration of nicotine in adult and adolescent mice Mice were chronically infused with nicotine at 48 mg/kg/day for 10 days via osmotic minipump. On day 11, mice were challenged with nicotine, and they were evaluated in three tests: tail-flick, hot-plate, and hypothermia. ED50 values (± CL) were calculated from the dose-response curve of the respective treatment, and they are expressed as milligrams per kilogram. Each dose group included 8 to 10 animals.
Potency ratios for tolerance studies Potency ratios with confidence intervals are given for each group. Adolescents showed a higher degree of tolerance to nicotine than adults in the hot-plate test.
Withdrawal following repeated injections in adult (PND 75) and adolescent (PND 28) mice. Male mice were injected with saline or 2.0 mg/kg nicotine s.c. three times a day for 4 days. Sixteen hours following the last injection, mice were injected with 2.0 mg/kg mecamylamine or saline s.c. and evaluated for somatic signs (a), elevated plus-maze (b), hot-plate (c), and locomotor activity (d). Each point represents the mean ± S.E. of 10 mice. *, p < 0.05 from saline group. MP, minipump.

Nicotine acute pharmacological effects in adult and adolescent mice. Mice from two age groups (PND 28 and PND 75) were injected s.c. with various acute doses of nicotine, and they were tested in the following responses: hypothermia (a), hot-plate test (b), tail-flick test (c), and locomotor activity (d). Each point represents the mean ± S.E. of 12 mice.
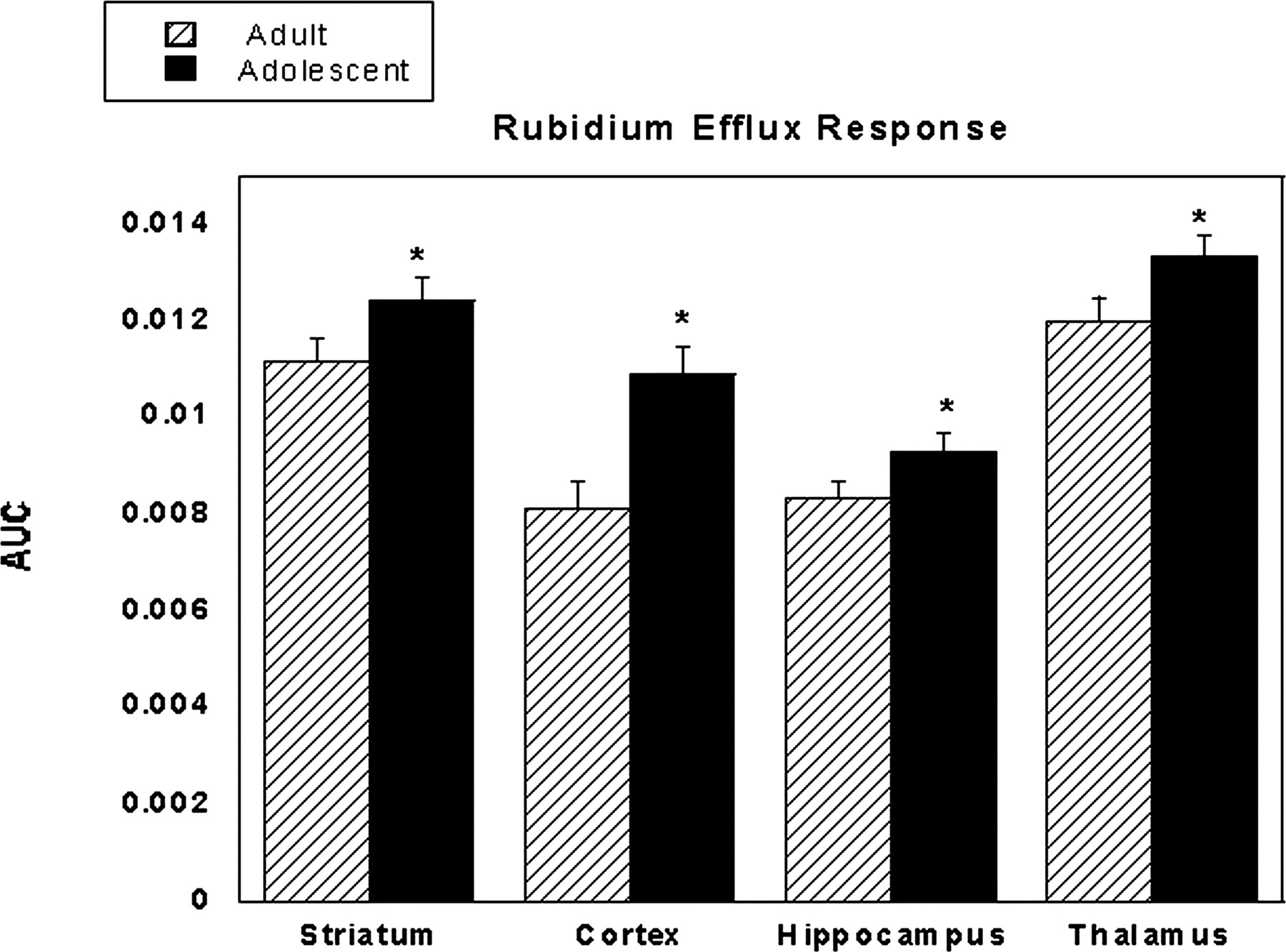
86Rb+ Efflux. Dose-response curves for nicotine-stimulated synaptosomes were generated for both adults and adolescents in four brain regions: striatum, cortex, hippocampus, and thalamus [examples shown in Fig. 7, and data were represented by a Michaelis-Menten curve fit where y = m1 × x/(m2 + x)]. Synaptosomes were responsive to nicotine stimulation in a dose-dependent manner until approximately 100 μM nicotine, at which point the response reached its peak. Adolescent mice displayed greater nAChR functionality with larger differences at lower concentrations of nicotine. The difference is also evident by the shift to the left of nicotine dose-response curves in adolescent compared with adult mice (Fig. 7). The increase in nicotine potency was also observed in the estimated EC50 values for both age groups in all brain areas tested. For adolescents values were striatum, 0.53 μM; cortex, 0.79 μM; hippocampus, 0.96 μM; and thalamus, 0.53 μM. EC50 values for adults were as follows: striatum, 1.61 μM; cortex, 10.77 μM; hippocampus, 3.13 μM; and thalamus, 3.93 μM. In addition, the determination of the total AUC showed that adolescent mice displayed significantly higher nicotinic receptor functionality than adults in all four brain regions tested (Fig. 8).
Discussion
The goal of this study was to investigate the differences seen across adult and adolescent rodents in two key components of nicotine dependence, namely, reward and withdrawal. Furthermore, initial sensitivity, tolerance, and in vitro receptor function were assessed to account for observed differences. We report here that adolescent male mice are more sensitive to the rewarding effects of nicotine than adult male mice in the conditioned place preference model. Several factors could explain the differences in nicotine potency. One such factor is differences in motor function between adult and adolescent mice. However, data from the locomotor activity tests report similar potency for both ages. Alternatively, the conditioning session and duration might have been optimal for adolescents but not for adults at a low dose of nicotine. Varying the duration of the sessions may alter the development of place preference at lower doses in adults and adolescents. However, previous studies have tested a variety of conditioning sessions and durations and have not identified this as a confounding factor (Belluzzi et al., 2004). An additional factor when using conditioned place preference involves contextual learning and memory. It is possible that adolescents display increased learning and make drug-context associations more rapidly than do adults. Indeed, adolescent rodents performed better in an active avoidance task (Spear and Brake, 1983). In addition, several reports found that adolescent rats displayed a significant conditioned place preference after only one pairing of nicotine, whereas adults showed no preference (Belluzzi et al., 2004; Brielmaier et al., 2007). Finally, it is possible that pharmacokinetic differences between adult and adolescent rodents could explain the different potencies. If this is the case, the adolescents are reported to have increased metabolism (Trauth et al., 2000); this, however, would translate into a decrease, not an increase, in the rewarding effect of nicotine. Furthermore, metabolism would not play a role in differences in the in vitro receptor potency. Given that metabolism is an unlikely contributing factor, our data suggest that pharmacological factors are responsible for the enhanced rewarding effect from nicotine conditioning in adolescent rodents.

Dose-response curve of nicotine after chronic administration in adult (PND 75) and adolescent (PND 28) mice. Animals were chronically infused with saline or nicotine at 48 mg/kg/day for 10 days via osmotic minipump. On day 11, mice were challenged with nicotine, and then they were evaluated in tail-flick and hot-plate analgesia and hypothermia. Adolescent mice are shown in a to c, and adult mice are shown in d to f. Each point represents the mean ± S.E. of 12 mice.
Data from our withdrawal study confirmed results found by O'Dell et al. (2006) in which somatic signs were attenuated in adolescent rats. In addition, our data also demonstrates that adolescent mice displayed decreased withdrawal signs in two other physical measures (hyperalgesia and hyperactivity) and one affective measure (anxiety-like behavior). However, results should be interpreted with caution because of several possible confounding factors. Although the use of minipumps allows for a minimal amount of stress to be placed on the animal, it will not take into account the fact that adolescent animals display a growth rate much faster than that of adults. Because the dose of nicotine is based on body weight at the beginning of the experiment, it could be argued that the younger mice are not receiving a dose of nicotine that is consistent with that of the adult. For this reason, we also evaluated withdrawal using a repeated injection model that was based on the daily weight of the animal. In both studies, it was found that withdrawal signs were less in adolescent than adult animals. Another factor that could be playing a role is that adults and adolescents could have differing sensitivities to mecamylamine. To investigate this possibility, we conducted a spontaneous withdrawal experiment in which mecamylamine was not used to precipitate withdrawal. Again, similar and consistent results were found in this study with adolescent animals displaying a decrease in the intensity of withdrawal signs. Collectively, these studies suggest a minor role for the two above-mentioned factors in the withdrawal differences. Overall, our data show that during adolescence, positive rewarding effects of nicotine are enhanced and negative withdrawal effects are less prominent. This characterization has the potential to contribute significantly to adolescent nicotine addiction. Indeed, if adolescents experience fewer aversive effects of nicotine and enhance pleasurable effects, they are more likely to smoke more often, which are behaviors critical in the progression to dependence.

Dose-response curves from the thalamus and cortex regions of adult (PND 75) and adolescent (PND 28) male mice. Synaptosomes from brain tissue were stimulated with various doses of nicotine for 1 min to generate dose-response curves. Area under the curve is shown on the y-axis, and nicotine dose is shown on the x-axis. Data are represented by a Michaelis-Menten curve fit where y = m1 × x/(m2 + x). In the thalamus, R values = 0.98 (adolescents) and 0.75 (adults). In the cortex, R values = 0.99 (adolescents) and 0.81 (adults). Adolescent mice (dashed line) displayed higher nAChR functionality compared with adult mice (solid line). Results are expressed as mean AUC ± S.E.

Total area under the curve for all doses of nicotine in the rubidium efflux assay in four brain regions. Results are expressed as mean AUC ± S.E. of 10 to 12 mice. *, p < 0.05 from adult mice.
Other mechanisms could underlie the differences seen in our behavioral observations. Functional properties, distribution, and number of nAChRs could differ between adults and adolescents. Indeed, our data suggest that there is either increased nAChR function or an increased quantity of brain nAChRs in adolescent mice. We again remind the reader that main comparisons are focused on total AUC. Although the estimated EC50 values do imply age differences in response to nicotine stimulation, further experimentation may be needed to refine the values. Comparisons of total AUC, however, do show clear and significant differences in nAChR functionality (Fig. 8). Indeed, the increased functionality of adolescent receptors could be playing a role in the behavioral observations seen in the conditioned place preference and withdrawal models. This work also agrees with our recent findings in the rat (Britton et al., 2007) where nicotine-stimulated rubidium efflux peaked during adolescence (∼PND 35). Recent research suggests that the mRNA expression and binding of some nAChR subunits, namely, α5, α6, and α7, is higher in early adolescence compared with adulthood in rats (Azam et al., 2007). In addition, regulation of nAChRs may vary after acute and chronic exposure to nicotine. It is possible that the enhanced rewarding response seen in the conditioned place preference model is due to increased nAChR functionality. However, it does not correlate with the data from our acute studies in which adolescent and adult mice displayed no significant differences in three of four measures in responses to acute nicotine. In contrast, a previous study in C57BL/6J mice found that adults were more affected by nicotine in both hypothermia and locomotor activity tests (Lopez et al., 2003). Certainly, strain and dosing differences could contribute to this discrepancy. Furthermore, it may be that differences in the biochemical measure are not enough to reach behavioral thresholds in a model using short exposure to nicotine such as that in our acute studies. However, a subchronic or chronic dosing protocol, such as that used in conditioned place preference, is enough to surpass this threshold, which is why differences are seen using this model.
Following acute treatment with nicotine, adolescent mice displayed an increased nicotine-induced antinociception compared with adults in the tail-flick test. This finding suggests that, in adolescents, predisposition to maintain use of nicotine might be due to the lessening of aversive effects due to decreased sensitivity to the drug.
The data from our tolerance study show that adolescent mice produce a greater degree of tolerance to nicotine-induced antinociception than adult mice. A prior report found that adolescent rats also showed tolerance to the locomotor depressant effects of nicotine at a higher dose compared with adults (Brielmaier et al., 2007). The tolerance level is an important factor in evaluating nicotine dependence. The higher tolerance seen in adolescents suggests that this age group would have to smoke more to achieve the same level of effect as an adult, leading to a greater intake of nicotine in adolescence and an increase in the likelihood of becoming dependent.
This study demonstrates that adult and adolescent animals do not exhibit aspects of nicotine dependence equally. Adolescent animals experience increased rewarding effects to nicotine as well as decreased withdrawal signs, which may lead to an overall shift in the vulnerability to nicotine addiction. It has been recognized that nicotine dependence rates are higher for adults who begin smoking during the adolescent period. Clearly, if adolescent smokers are experiencing enhanced rewarding properties and attenuated aversive properties, they will be more likely to continue smoking behavior.
Taken together, this study has important implications for the mechanism of nicotine dependence in adolescence. The decreased initial sensitivity and increased tolerance to nicotine may contribute to an enhanced vulnerability. In addition, the increase in adolescent nAChR functionality compared with adults may contribute to the shift in nicotine dependence. Because neuronal pathways are still developing in young animals, it is possible that the adaptations in the brain during development cause the levels of dependence to change over time as well. For example, the dopaminergic system is under great development during adolescence. Azam et al. (2007) report that nicotine-stimulated dopamine release is significantly higher during the early adolescent period in the male rat. Because dopamine plays a large role in the rewarding effects of nicotine, this observation may explain observations from our conditioned place preference model. Furthermore, previous work has demonstrated that dopamine release is attenuated in the adult rat during withdrawal (Hildebrand et al., 1998; Di Chiara, 2000). Therefore, it is possible that adolescents do not experience this same decrease in dopamine, thus lowering withdrawal symptoms and aversive effects.
These data indicate that the underlying mechanisms of nicotine dependence differ for adults and adolescents, suggesting that the effectiveness of smoking cessation therapies should not be considered equal for all groups. For example, smoking cessation therapies aimed at minimizing withdrawal effects may not be as effective in adolescents who do not experience these symptoms in the same manner as adults. In conclusion, this study indicates that adolescence should be considered a critical timeframe for the ability to acquire nicotine dependence.
Acknowledgments
We greatly appreciate the technical assistance of Tie Han.
Footnotes
-
This study was supported by Virginia Tobacco Settlement Foundation through the Virginia Youth Tobacco Project to Virginia Commonwealth University.
-
Article, publication date, and citation information can be found at http://jpet.aspetjournals.org.
-
doi:10.1124/jpet.107.121616.
-
ABBREVIATIONS: PND, postnatal day; nAChR, nicotinic acetylcholine receptor; %MPE, percent maximal possible effect; ANOVA, analysis of variance; CL, confidence limits; AUC, area under the curve.
- Received February 16, 2007.
- Accepted April 18, 2007.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}